



研究内容
酒粕は色白がお好み?麹菌がつくるチロシナーゼを発見

アミノ酸のチロシンを酸化してメラニンを生じさせるチロシナーゼに関する研究は、ヒトでは肌の色を白くする目的で、阻害剤をはじめとする研究が幅広く行われています。また、この酵素は微生物から哺乳類まで幅広く存在し、植物では生体防御に関っているとも言われています。
月桂冠総合研究所では、麹菌Aspergillus oryzaeから固体培養(米麹など)で特異的に発現するチロシナーゼを発見しました。これまでは、微生物由来のチロシナーゼに関する報告は多くありませんでした。本研究は、さらにチロシナーゼが特定の条件で発現することを明らかにしたことから、チロシナーゼの役割を研究する上で非常に興味深いものであると考えています。
研究テーマ 麹菌の固体培養特異的に発現するチロシナーゼのクローニングと大量発現
実験目的
清酒業界では昭和30年ごろ、酒粕に黒い斑点が生じる「黒粕(くろかす)」という現象に悩まされました。酒粕は食品として幅広く利用されており、この問題を解決するために、多くの精力的な研究が行われました。その結果、酒粕中に含まれるアミノ酸(チロシン)が酸化されて生成するメラニンが黒い斑点の原因であること、この酸化反応には麹菌のチロシナーゼという酵素が関与していること、が明らかとなりました1)(現在ではチロシナーゼ活性の低い麹菌の育種が進み「黒粕」の発生を未然に防ぐことができています)。
一方、近年の遺伝子解析技術の驚異的な発達により、細菌(大腸菌、納豆菌、乳酸菌など)をはじめ、真核微生物(酵母など)、ついにはヒトの全遺伝子配列が明らかとなりました。当社は産学官の麹菌遺伝子解析プロジェクトに参画し2)、ここで得た情報と独自の研究により得た知見から「黒粕」の原因となる新規チロシナーゼの遺伝子を単離し、麹菌で大量に発現させることに成功しました。
実験結果および考察
麹菌のチロシナーゼ遺伝子は1995年に、一島ら3)により初めて単離されmelO遺伝子と名付けられましたが、我々はこの遺伝子が液体培養でしか発現しないことを明らかにしました4)。このことから、酒粕の褐変には固体培養でのみ発現する未知のチロシナーゼが関与していると考えられました。
そこで、麹菌の遺伝子発現データベースを詳細に解析した結果、液体培養のチロシナーゼに少し似た遺伝子が固体培養のデータに含まれていることを見出しました5)。我々はこの遺伝子が固体培養でのみ発現しているかどうかを液体培養菌体と米麹菌体で確認する実験をノザン解析により行いました。
その結果、この遺伝子は米麹で特異的に発現しており、液体培養では全くシグナルが観察されませんでした(図1)。このことから、この遺伝子は明らかに固体培養特異的に発現していることが分かりました。
この遺伝子がチロシナーゼをコードすることを確認するため、配列を確認したところチロシナーゼの活性に必要とされる領域が確かに存在することが分かりました。なお、一般的にチロシナーゼは活性に必要な領域以外はあまり似ておらず、この遺伝子がどのように進化してきたのか興味深い点です。

今回発見したチロシナーゼが「黒粕」の原因となる麹の褐変に関与しているかどうか確認するため、麹菌での高発現を試みました。melB遺伝子のコード領域を、米麹で非常に強力に働くことが知られているglaBプロモーター支配下で高発現させたところ、親株に比べて非常に強い褐変性を示しました(図2)。

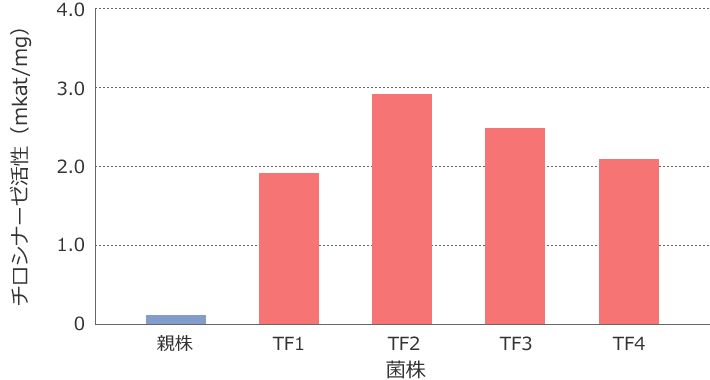
また、チロシナーゼ活性を測定したところ形質転換株で作成した麹は親株の10倍以上のチロシナーゼ活性を示したことから、この遺伝子は確かにチロシナーゼをコードしており、この酵素が米麹の褐変(「黒粕」)の原因であることが明らかとなりました6)(図3)。
これまでに当社研究所では麹菌がglaAグルコアミラーゼ(液体培養特異的)とglaBグルコアミラーゼ(固体培養特異的)の二つを培養条件によって遺伝子の発現を巧みに使い分けていることを明らかにしました7)。さらに今回新たに、麹菌がmelOチロシナーゼ(液体培養特異的)とmelBチロシナーゼ(固体培養特異的)の遺伝子の発現を使い分けている事実が明らかとなりました。このように環境条件による遺伝子発現の使い分けは、麹菌の進化のどの過程で獲得されたものか非常に興味深く、今後はmelO、melBのタンパク質の性質の違いを研究することにより、その謎を追求していきたいと考えます。
近年の研究では、melBはglaBと同じように固体培養特異的に発現するだけでなく、さらに培養時間が長くなるほど発現強度が高くなることが報告されています(岩手大学の研究グループ)8)。この結果は、米麹の製造期間を長くすると黒粕を生じやすいといった知見と一致することがわかりました9)。またこのチロシナーゼは、天然由来の髪色素を作るのにも用いられ、醸造産業以外にも活用されています(図4)。
学会発表
- 新規チロシナーゼ遺伝子(melB)と麹褐変性との関係、醸造学会(2002)
○中村幸宏、小畑浩、秦洋二、川戸章嗣、安部康久 - 麹菌の固体培養特異的に発現するチロシナーゼ遺伝子のクローニングと機能解析、真核微生物交流会(2001)
小畑浩 - 麹菌の固体培養特異的に発現するチロシナーゼ遺伝子のクローニングと機能解析、農芸化学会(2001)
○小畑浩、石田博樹、秦洋二、川戸章嗣、杉並孝二、安部康久、一島英治 - Two tyrosinase-encoding genes in Aspergillus oryzae - culture-specific gene expression -、国際糸状菌分子生物学コンファレンス(2000)
○小畑浩、石田博樹、秦洋二、川戸章嗣、安部康久、赤尾健、秋田修、一島英治 - 麹菌の固体培養特異的に発現する新規チロシナーゼ遺伝子のクローニングと解析、生物工学会(2000)
○小畑浩、石田博樹、秦洋二、川戸章嗣、安部康久、赤尾健、秋田修、一島英治
論文
- Cloning of a Novel Tyrosinase-Encoding Gene(melB) from Aspergillus oryzae and Its Overexpression in Solid-State Culture(Rice Koji)
H. Obata, H. Ishida, Y. Hata, A. Kawato, Y. Abe, T. Akao, O. Akita, E. Ichishima : J. Biosci. Bioeng., 97, 400-405 (2004)
参考文献
- T. Ohba, H. Kato, T. Kurata, M. Fujimaki.: Agr. Biol. Chem., 39, 139 (1975)
- 町田雅之;化学と生物, 39, 349 (2001)
- Y. Fujita, Y. Uraga, and E. Ichishima : Biochim. Biophys. Acta., 1261, 151 (1995)
- H. Ishida, K. Matsumura, Y. Hata, A. Kawato, K. Suginami, Y. Abe, S. Imayasu, E. Ichishima: Appl Microbiol Biotechnol., 57, 131(2001)
- 小畑浩ら;平成12年度生物工学会大会講演要旨集、p19
- H. Obata, H. Ishida, Y. Hata, A. Kawato, Y. Abe, T. Akao, O. Akita, E. Ichishima : J. Biosci. Bioeng., 97, 400-405 (2004)
- H. Ishida, Y. Hata, E. Ichikawa, A. Kawato, K. Suginami, S. Imayasu :J. Ferment. Bioeng., 86, 301-307 (1998)
- J. Chen, A. Tonouchi :Biosci. Biotechnol. Biochem., 88, p220–224 (2024)
- 大場, 日本醸造協会誌, 66, p864-869 (1971)
(掲載日:2004年9月1日)
(改訂日:2025年6月5日)



